Моя статья о работе энергетических систем будет полезной тем, кто ходит на семинары инструкторов тренажёрного зала или групповых программ. Многие эту тему не понимают или понимают неправильно. Ниже я вкратце попытался разъяснить принцип включения различных систем в энергообеспечение физических нагрузок.
Д.А.Жабкин
РАБОТА ЭНЕРГЕТИЧЕСКИХ СИСТЕМ ОРГАНИЗМА ПРИ АЭРОБНЫХ ФИЗИЧЕСКИХ НАГРУЗКАХ
Общая характеристика энергетических систем организма
Для любого физиологического процесса в организме, требуется энергия. При мышечной деятельности происходит процесс преобразования химической энергии в механическую работу. Универсальным источником энергии в живом организме является молекула АТФ. Под действием фермента Ca2+-АТФ-азы АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты, и превращается в АДФ, при этом высвобождается энергия.
АТФ + H2O → АДФ + H3PO4 + 7,3 ккал (или 30 кДж)
Запас молекул АТФ в мышце ограничен (около 5 ммоль*кг-1 сырой массы ткани), что может обеспечить выполнение интенсивной работы в течение очень короткого времени (0,5-1,5 секунды или 3-4 одиночных сокращения максимальной силы). Поэтому расход энергии при работе мышцы требует постоянного его восполнения.
Дальнейшая мышечная работа происходит благодаря быстрому ресинтезу АТФ из продуктов её распада и такого количества энергии, которое выделилось при распаде:
АДФ + H3PO4 + 7,3 ккал → АТФ
Мышца имеет 3 основных источника воспроизводства энергии:
1. Богатые энергией фосфатосодержащие вещества, которые присутствуют в тканях (АДФ, креатинфосфат);
2. Богатые энергией фосфатосодержащие вещества, которые образуются в процессе катаболизма гликогена, жирных кислот и других энергетических субстратов (дифосфоглицериновая кислота, фосфопировиноградная кислота и др.);
3. Энергия протонного градиента на мембране митохондрий, образующаяся в результате окисления различных веществ.
В зависимости от того, с помощью какого биохимического процесса поставляется энергия для получения молекул АТФ, выделяют 4 механизма ресинтеза АТФ в тканях или энергетические системы организма.
Для того, чтобы понять основные отличия энергетических систем, пользуются следующими характеристиками:
Ёмкость энергетической системы – это количество АТФ, способное образоваться за счёт данной системы.
Мощность энергетической системы – это количество АТФ, производимое системой за единицу времени.
Скорость развёртывания – время достижения максимальной мощности системы от начала работы.
Метаболическая эффективность – та часть энергии, которая накапливается в макроэргических связях АТФ. Она определяет экономичность выполняемой работы и оценивается коэффициентом полезного действия.
Таблица 1. Общая характеристика энергетических систем
| система | мощность, дж*кг*мин-1 | максимальная мощность | ёмкость, кДж*кг-1 | субстраты | основное ограничение | существенная роль | время восстановления |
|---|---|---|---|---|---|---|---|
| Фосфогенная | 3770 | 6-12 секунд, Время развёртывания: 0,5-0,7 сек | 630 | АТФ, КФ | содержание КФ | интенсивная кратковременная работа 2-30 сек | 40-60 мин |
| Лактатная | 2500 | 60-180 секунд Время развёртывания: через 20-40 сек | 1050 | глюкоза, гликоген | накопление молочной кислоты | кратковременная интенсивная работа от 3 сек до 3-х минут | 2-5 час |
| Аэробная | 1250 | 6-10 минут. Время развёртывания: через 2-3 мин | ∞ | глюкоза, гликоген | количество гликогена, скорость доставки О2 | низкоинтенсивная аэробная нагрузка до 20 минут | 5-24 час |
| жирные кислоты | скорость доставки О2 | низкоинтенсивная аэробная нагрузка более 20 минут | Сутки, несколько суток |
Данные в таблице 1 получены путём измерения данных показателей у высококвалифицированных спортсменов. У нетренированных людей данные значения ниже.
Теперь остановимся поподробнее на отдельных энергетических системах.
Креатинфосфатная (фосфогенная, алактатная) система
АТФ в этой системе образуется в результате реакции Ломана, которая происходит в присутствии фермента креатинфосфаткиназы.
АДФ + КреатинФосфат → АТФ + креатин
Запасы креатинфосфата в волокне в 3-4 раза выше, чем АТФ. Но этого количества хватает для использования его в качестве источника энергии только на начальном этапе работы мышцы в первую минуту, до момента активизации других более мощных источников. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Эта система определяет алактатную работоспособность мышц.
Максимальная алактатная мощность зависит от:
1. концентрации и активности фермента креатинфосфаткиназа (переносящего фосфатную группу с креатинфостфата на АДФ).
2. концентрации креатинфосфата.
Длительность удержания максимальной алактатной мощности составляет 6-12 секунд.
Алактатная емкость зависит от запасов креатинфосфата в мышце.
Эффективность креатинфосфаткиназной реакции очень велика (76%), так как реакция протекает непосредственно между двумя веществами на миофибриллах.
Лактатная (гликолитическая, лактацидная) система
Гликолиз – это процесс распада одной молекулы глюкозы на две молекулы молочной кислоты с выделением энергии, достаточной для фосфорилирования двух молекул АТФ, протекает в саркоплазме под воздействием 10 ферментов.
C6H12O6 + 2H3PO4 + 2АДФ → 2C3H6O3 + 2АТФ + 2H2O
Гликогенолиз – это процесс распада гликогена.
[C6H10O5] n + 3H3PO4 + 3АДФ → 2C3H6O3 + [C6H10O5] n-1 + 3АТФ + 2H2O
Для работы этой системы используются в основном внутримышечные запасы гликогена, а также глюкоза, поступающая из крови.
Гликолиз протекает без потребления кислорода и способен быстро восстанавливать запасы АТФ в мышце. Достигает максимума через 30-40 секунд интенсивной работы.
Эта система определяет лактатную работоспособность мышц.
Максимальная лактатная мощность определяется главным образом концентрацией и активностью ключевых ферментов гликолиза, которые зависят от:
1. устойчивости ферментов гликолиза к повышению кислотности среды, которая ингибирует их активность.
2. устойчивости кислотно-щелочного равновесия внутренней среды мышц, в условиях усиленной выработки молочной кислоты.
Время удержания максимальной мощности данного метаболического процесса составляет 60-180 секунд.
Гликолитическая емкость определяется главным образом запасами гликогена в мышцах, гликоген печени для процессов гликолиза не обладает достаточной мобильностью.
Метаболическая эффективность гликолиза оценивается значениями КПД порядка 0,35-0,52. Это означает, что почти половина всей выделяемой энергии превращается в тепло и не может быть использована в работе.
Умеренный сдвиг pH в кислую сторону активирует работу ферментов дыхательного цикла в митохондриях и усиливает аэробное энергообразование.
Значительное накопление молочной кислоты, появление избыточного СО2, изменение рН и гипервентиляция лёгких, отражающие усиление гликолиза в мышцах, обнаруживаются при увеличении интенсивности нагрузки более 50% максимальной аэробной мощности. Этот уровень нагрузки обозначается, как порог анаэробного обмена (ПАНО). Чем раньше он будет достигнут, тем быстрее вступит в действие гликолиз, сопровождающийся накоплением молочной кислоты и последующим развитием утомления работающих мышц.
Величина ПАНО является важным показателем эффективности энергообразования в мышцах, роста степени тренированности, который широко используется при биохимическом контроле функционального состояния спортсмена. С ростом степени тренированности на выносливость ПАНО увеличивается, т.е. наступает при более интенсивной работе.
Миокиназная реакция
«Аварийный» путь ресинтеза АТФ:
АДФ + АДФ → АТФ + АМФ
Происходит в мышцах при значительном увеличении концентрации АДФ в саркоплазме. Такая ситуация возникает при выраженном мышечном утомлении, когда другие пути ресинтеза АТФ уже не справляются.
Эта реакция так же обратима и используется для поддержания постоянного уровня АТФ в мышцах.
Аэробная (кислородная, окислительная) система.
В обычных условиях аэробный механизм ресинтеза АТФ обеспечивает около 90% общего количества АТФ, ресинтезируемой в организме.
Окисление протекает в митохондриях под воздействием специальных ферментов и требует затрат кислорода, а соответственно и времени на его доставку. Такие процессы называются аэробными. Окисление происходит в несколько этапов, сначала идет гликолиз (см. выше), но образовавшиеся в ходе промежуточного этапа этой реакции две молекулы пирувата не преобразуются в молекулы молочной кислоты, а проникают в митохондрии, где окисляются в цикле лимонной кислоты до углекислого газа и воды, давая энергию для производства еще 36 молекул АТФ.
C6H12O6 + 6O2 + 38АДФ + 38H3PO4 → 6CO2 + 44H2О + 38АТФ
Итого распад глюкозы по аэробному пути дает энергию для восстановления 38 молекул АТФ. Т.е. окисление в 19 раз эффективнее гликолиза. Если во время гликолиза организм усваивает в виде АТФ лишь 3% энергии, заложенной в молекуле глюкозы, то при аэробном окислении этот показатель равен 55% (включая те самые 3%). К тому же аэробное окисление может использовать более энергоемкие субстраты, такие как жиры, которые дают в 2 раза больше энергии, чем то же количество углеводов.
Субстратами окисления являются любые органические вещества: белки, жиры, углеводы. Долевое участие будет зависеть от характера работы:
| Интенсивность нагрузки | Преимущественно используются |
|---|---|
| до 50% МПК (лёгкая работа) | жиры |
| 60-90% МПК (тяжёлая работа) | углеводы |
| близкая к МПК | практически только углеводы |
Эта система определяет аэробную работоспособность мышц.
Максимальная аэробная мощность зависит главным образом от:
1. плотности митохондрий в мышечных волокнах;
2. концентрации и активности окислительных ферментов;
3. скорости поступления кислорода вглубь волокна.
Объем кислорода доступного для окислительных реакций лимитируется:
1. состоянием кардио-респираторной системы;
2. капилляризация мышц;
3. концентрация миоглобина;
4. диаметр мышечного волокна (чем меньше диаметр волокна, тем лучше оно снабжается кислородом и тем выше его относительная аэробная мощность).
Показатель количества кислорода, усваиваемого единицей массы тела за единицу времени – МПК (максимальное потребление кислорода).
Скорость производства АТФ за счет окисления достигает максимальных значений на 2-3-й минуте работы, что связано с необходимостью развертывания множества процессов, обеспечивающих доставку кислорода к митохондриям. Время удержания максимальной аэробной мощности составляет примерно 6 минут, в дальнейшем аэробная мощность снижается по причине усталости всех активно работающих систем организма.
Аэробная ёмкость очень высокая, т.к. для окисления используются любые органические вещества.
Метаболическая эффективность этого механизма – около 50%. Она определяется по ПАНО: у нетренированных людей ПАНО наступает при потреблении кислорода примерно 50% от МПК, а у высокотренированных на выносливость – при 80-90% от МПК.
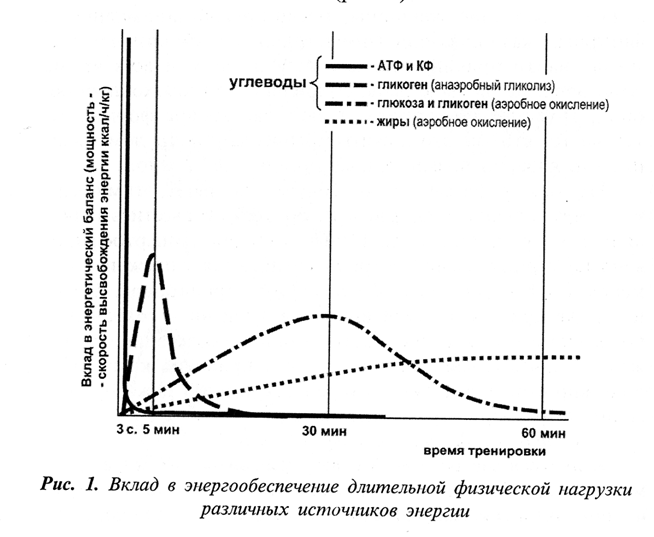
Общая характеристика аэробных физических нагрузок
В спортивный практике к упражнениям аэробного характера относят длительные физические упражнения, где относительный вклад аэробного процесса в затратах энергии превышает 70%.
К аэробным циклическим упражнениям (по Я.М.Коцу, 1986) относятся:
1. упражнения максимальной аэробной мощности (3-10 минут);
2. упражнения близкой к максимальной аэробной мощности (10-30 минут);
3. упражнения субмаксимальной аэробной мощности (30-80 минут);
4. упражнения средней аэробной мощности (80-120 минут);
5. упражнения малой аэробной мощности (более 120 минут).
Аэробный механизм является основным при таких видах спорта, как: бег на дистанции 5-25 км, велогонки, плавание на 800-1500 м, бег на коньках на 5-10 км и др.
Ёмкость аэробного механизма, которая в значительной степени определяется запасами гликогена в скелетных мышцах и печени, а также уровнем утилизации кислорода мышцами, существенно повышается уже в течение 1,5-2 месяцев тренировки на выносливость.
Мощность аэробного механизма, которая зависит от МПК и активности окислительных ферментов, также увеличивается в процессе адаптации к мышечной деятельности через 2-3 месяца тренировок.
Аэробная направленность физических нагрузок происходит, как правило, в зоне умеренной мощности. При этом упражнения выполняются при максимуме аэробного производства энергии.
Кислородный запрос может достигать 500-1500 л, кислородный долг не превышает 5 л (до 10%). Содержание молочной кислоты в крови составляет 0,6-0,8 г*л-1. В ходе работы она может извлекаться тканями и аэробно окисляться в них.
Вследствие усиленного использования запасов гликогена в печени, содержание глюкозы в крови становится ниже 0,8 г*л-1. В моче в значительном количестве появляются продукты распада белков. Отмечается потеря организмом воды и минеральных солей.
Основными упражнениями для развития аэробных процессов энергообеспечения будут физические нагрузки, относящиеся к зоне большой и умеренной мощности с интенсивностью работы на уровне ПАНО и 100% МПК.
Работа энергетических систем во время аэробной физической нагрузки
Для большей вариабельности рассмотрим несколько вариантов аэробной физической нагрузки.
При беге на длинные дистанции (5 и 10 км) аэробное окисление углеводов является основным механизмом энергообеспечения работы, так как на его долю приходится до 87% общих затрат энергии на дистанции 5 км и 97% на дистанции 10 км.
Вклад анаэробных источников на этих дистанциях также достаточно большой. Он может достигать 15% общих затрат энергии и играет важную роль при финишном ускорении, приносящем победу при беге на длинные дистанции.
Наиболее значительным фактором, влияющим на выносливость, является кислородное снабжение работающих мышц, поскольку потребление кислорода во время бега поддерживает максимальную скорость окисления углеводов. Порог анаэробного обмена у стайеров при работе достигается при 75-90% МПК.
При марафонском беге затраты энергии восполняются исключительно за счёт аэробного процесса. Погашение этих затрат невозможно только за счёт окисления углеводов из-за недостаточности запасов гликогена в работающих мышцах спортсмена, поэтому значительная часть энергии образуется за счёт окисления жиров, на долю которых может приходится от 10 до 50% общих затрат энергии.
Вклад жиров на длинных и сверхдлинных дистанциях у высокотренированных бегунов с большими запасами гликогена в работающих мышцах составляет 12-20%, у нетренированных бегунов – более 80%. Всего на дистанции марафонского бега окисляется около 300 г жиров.
Использование жиров в качестве источника энергии менее эффективно по сравнению с окислением углеводов, так как происходит с более низкой скоростью и с большим потреблением кислорода.

Рис 2. Механизмы энергообеспечения бега на 10 000 м и марафонского бега (пунктирная черта показывает момент исчерпания запасов гликогена)
При длительной работе наряду с увеличением использования в энергетическом обмене жиров может происходить новообразование углеводов из веществ неуглеводной природы (глюконеогенез).
Основным субстратом глюконеогенеза являются аминокислоты, часть которых накапливается в мышце при работе в результате распада тканевых белков.
Рассмотрим, как включаются в работу энергетические системы во время аэробных физических нагрузок.
Таблица 2. Вклад различных источников энергии в обеспечение ресинтеза АТФ при беге в аэробном режиме работы (в %).
| Дистанция, м | Креатинфосфат | Анаэробное окисление гликогена мышц | Аэробное окисление гликогена мышц | Глюкоза крови (гликоген печени) | Жирные кислоты |
|---|---|---|---|---|---|
| 1500 | Минимальный | 25 | 75 | — | — |
| 5000 | Минимальный | 12,5 | 87,5 | — | — |
| 10000 | Минимальный | 3 | 97 | — | — |
| Марафон | — | — | 75 | 5 | 20 |
| Супермарафон (84 км) | — | — | 35 | 5 | 60 |
| 24-часовой забег | — | — | 10 | 2 | 88 |
При переходе из состояния покоя к мышечной деятельности потребности в кислороде возрастает в несколько раз, но сразу она не может быть удовлетворена. Необходимо время, чтобы усилилась деятельность кардиореспираторной системы, чтобы кровь, обогащённая кислородом смогла дойти до работающих мышц. По мере усиления активности работы этих систем постепенно увеличивается потребление кислорода в работающих мышцах. Скорость потребления кислорода увеличивается до тех пор, пока не наступит истинное устойчивое состояние метаболических процессов, при котором потребление кислорода в данный момент времени точно соответствует потребности организма в нём (кислородному запросу).
До этого момента потребность организма в энергии обеспечивается большей частью за счёт работы анаэробных энергетических систем. Как мы уже отмечали выше, скорость развёртывания креатинфосфатной системы до полной мощности – доли секунды, лактатной – около половины минуты. В зависимости от того, какой кислородный запрос работы имеет нагрузка, кислородный дефицит на начальном её этапе восполняется за счёт разного участия анаэробных систем, но в любом случае развёртывания этих систем на полную мощность при нагрузках аэробного характера не требуется. В результате происходит накопление в организме недоокисленных продуктов анаэробного распада.
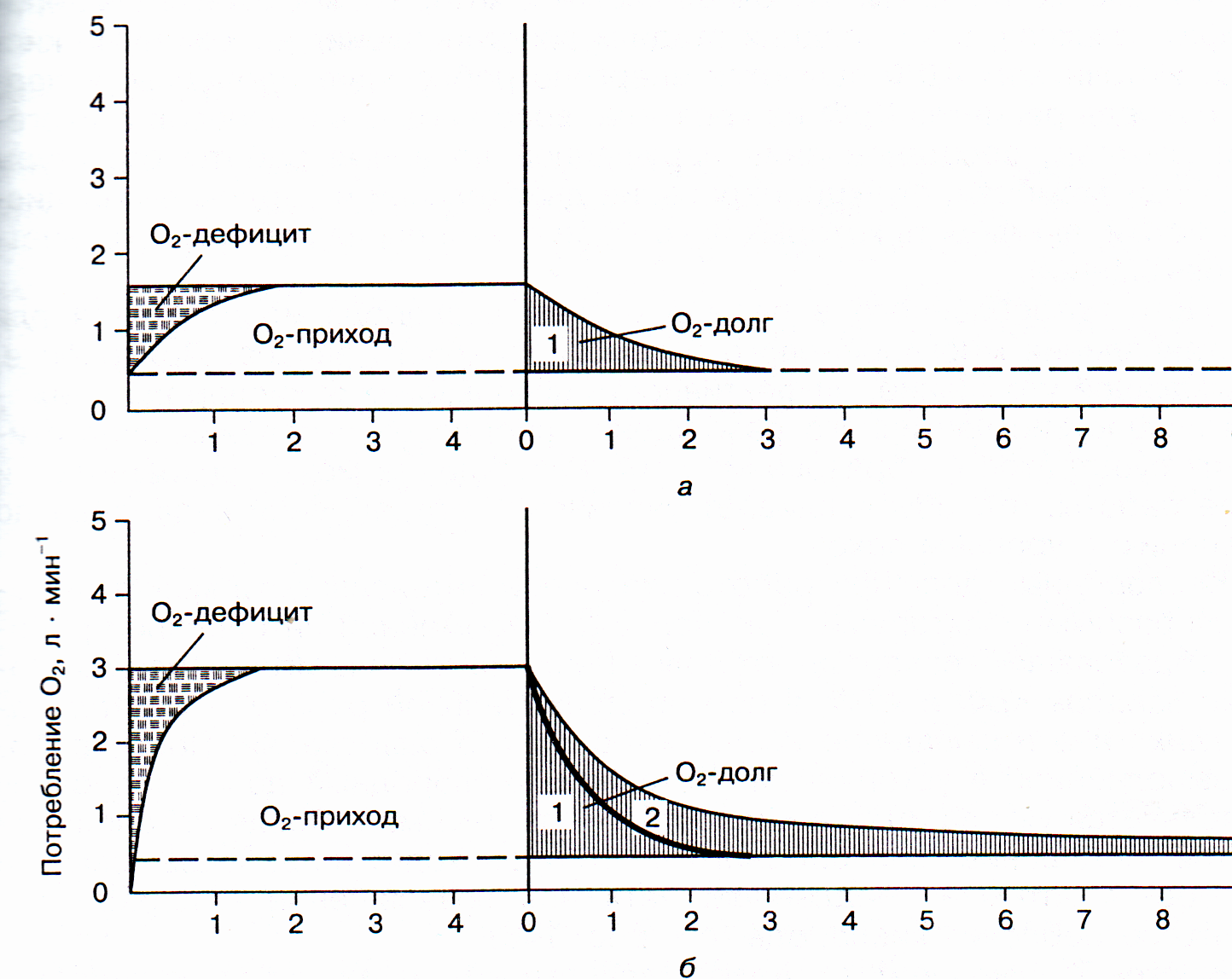
Рис 3. Кислородный приход, кислородный дефицит и кислородный долг при аэробной работе (а) лёгкой, (б) тяжёлой интенсивности. 1 – быстрый, 2 – медленный компоненты кислородного долга.
При работе в устойчивом состоянии часть анаэробных метаболитов может окисляться за счёт усиления аэробных реакций в процессе работы, а другая их часть устраняется после работы.
При выполнении работы с уровнем запроса около 50% МПК прирост концентрации молочной кислоты невелик (до 0,4-0,5 г/л), а при выполнении продолжительных нагрузок с уровнем запроса 50-85% МПК, возрастает до 1-1,5 г/л. Концентрация молочной кислоты значительно возрастает в первые 2-10 минут работы, а затем либо остаётся на прежнем уровне, либо снижается. То есть максимальная концентрация молочной кислоты в крови наблюдается до тех пор, пока не установилось устойчивое состояние, создающее условия для аэробного её окисления.
Для восстановления энергетических источников и окисления недоокисленных продуктов требуется дополнительное количество кислорода, поэтому некоторое время после окончания работы потребление его продолжает оставаться повышенным по сравнению с уровнем покоя. Этот излишек потребления кислорода в период восстановления получил название «кислородный долг».
Кислородный долг всегда больше кислородного дефицита. Чем больше интенсивность и продолжительность работы, тем кислородный долг выше.
После работы в устойчивом состоянии кислородный долг наполовину восполняется уже за 30 секунд, а полностью через 3-5 минут. После интенсивной работы «погашение» долга происходит в две фазы.
Быстрый (алактатный) компонент кислородного долга включает то количество кислорода, которое необходимо для ресинтеза АТФ и креатинфосфота. Он характеризует вклад креатинфосфатной энергетической системы в обеспечении выполненной работы.
Медленный (лактатный) компонент кислородного долга включает то количество кислорода, которое необходимо для окисления образовавшейся молочной кислоты при выполнении работы. Его величина характеризует участие лактатной энергетической системы, а при длительной работе – и других процессов, долю которых оценить весьма затруднительно. Медленный компонент устраняется наполовину за 15-25 минут, а полностью – за 1,5-2 часа.
Подводя итог, хочется отметить следующее:
• во время аэробной физической нагрузки работают все энергетические системы организма, но подавляющую роль играет аэробная система;
• все системы начинают работать одновременно с началом нагрузки, но за счёт разной скорости развёртывания процессов энергообразования, аэробная система полностью обеспечивает кислородный запрос не сразу, и на начальном этапе (несколько минут) кислородный приход компенсируют анаэробные энергетические системы.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности – Киев: Олимпийская литература, 2000
2. Граевская Н.Д., Долматова Т.И. Спортивная медицина: Курс лекций и практические занятия. В 2х частях. – М.: Советский спорт, 2004
3. Ким Н.К., Дьяконов М.Б. Фитнес. Учебник – М.: Советский спорт, 2006
4. Макарова Г.А. Медицинский справочник тренера – М.: Советский спорт, 2004
5. Руненко С.Д. Фитнес: мифы, иллюзии, реальность – М.: Советский спорт, 2005
6. Протасенко В.А. Думай! Или ‘Супертренинг’ без заблуждений, — журнал «Мускуляр», 2001
7. Хоули Э., Дон Френкс Б. Руководство инструктора оздоровительного фитнеса – Киев. Олимпийская литература, 2004
(С)2011 Д.А.Жабкин
Моя статья о работе энергетических систем будет полезной тем, кто ходит на семинары инструкторов тренажёрного зала или групповых программ. Многие эту тему не понимают или понимают неправильно. Ниже я вкратце попытался разъяснить принцип включения различных систем в энергообеспечение физических нагрузок.
Д.А.Жабкин
РАБОТА ЭНЕРГЕТИЧЕСКИХ СИСТЕМ ОРГАНИЗМА ПРИ АЭРОБНЫХ ФИЗИЧЕСКИХ НАГРУЗКАХ
Общая характеристика энергетических систем организма
Для любого физиологического процесса в организме, требуется энергия. При мышечной деятельности происходит процесс преобразования химической энергии в механическую работу. Универсальным источником энергии в живом организме является молекула АТФ. Под действием фермента Ca2+-АТФ-азы АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты, и превращается в АДФ, при этом высвобождается энергия.
АТФ + H2O → АДФ + H3PO4 + 7,3 ккал (или 30 кДж)
Запас молекул АТФ в мышце ограничен (около 5 ммоль*кг-1 сырой массы ткани), что может обеспечить выполнение интенсивной работы в течение очень короткого времени (0,5-1,5 секунды или 3-4 одиночных сокращения максимальной силы). Поэтому расход энергии при работе мышцы требует постоянного его восполнения.
Дальнейшая мышечная работа происходит благодаря быстрому ресинтезу АТФ из продуктов её распада и такого количества энергии, которое выделилось при распаде:
АДФ + H3PO4 + 7,3 ккал → АТФ
Мышца имеет 3 основных источника воспроизводства энергии:
1. Богатые энергией фосфатосодержащие вещества, которые присутствуют в тканях (АДФ, креатинфосфат);
2. Богатые энергией фосфатосодержащие вещества, которые образуются в процессе катаболизма гликогена, жирных кислот и других энергетических субстратов (дифосфоглицериновая кислота, фосфопировиноградная кислота и др.);
3. Энергия протонного градиента на мембране митохондрий, образующаяся в результате окисления различных веществ.
В зависимости от того, с помощью какого биохимического процесса поставляется энергия для получения молекул АТФ, выделяют 4 механизма ресинтеза АТФ в тканях или энергетические системы организма.
Для того, чтобы понять основные отличия энергетических систем, пользуются следующими характеристиками:
Ёмкость энергетической системы – это количество АТФ, способное образоваться за счёт данной системы.
Мощность энергетической системы – это количество АТФ, производимое системой за единицу времени.
Скорость развёртывания – время достижения максимальной мощности системы от начала работы.
Метаболическая эффективность – та часть энергии, которая накапливается в макроэргических связях АТФ. Она определяет экономичность выполняемой работы и оценивается коэффициентом полезного действия.
Таблица 1. Общая характеристика энергетических систем
| система | мощность, дж*кг*мин-1 | максимальная мощность | ёмкость, кДж*кг-1 | субстраты | основное ограничение | существенная роль | время восстановления |
|---|---|---|---|---|---|---|---|
| Фосфогенная | 3770 | 6-12 секунд, Время развёртывания: 0,5-0,7 сек | 630 | АТФ, КФ | содержание КФ | интенсивная кратковременная работа 2-30 сек | 40-60 мин |
| Лактатная | 2500 | 60-180 секунд Время развёртывания: через 20-40 сек | 1050 | глюкоза, гликоген | накопление молочной кислоты | кратковременная интенсивная работа от 3 сек до 3-х минут | 2-5 час |
| Аэробная | 1250 | 6-10 минут. Время развёртывания: через 2-3 мин | ∞ | глюкоза, гликоген | количество гликогена, скорость доставки О2 | низкоинтенсивная аэробная нагрузка до 20 минут | 5-24 час |
| жирные кислоты | скорость доставки О2 | низкоинтенсивная аэробная нагрузка более 20 минут | Сутки, несколько суток |
Данные в таблице 1 получены путём измерения данных показателей у высококвалифицированных спортсменов. У нетренированных людей данные значения ниже.
Теперь остановимся поподробнее на отдельных энергетических системах.
Креатинфосфатная (фосфогенная, алактатная) система
АТФ в этой системе образуется в результате реакции Ломана, которая происходит в присутствии фермента креатинфосфаткиназы.
АДФ + КреатинФосфат → АТФ + креатин
Запасы креатинфосфата в волокне в 3-4 раза выше, чем АТФ. Но этого количества хватает для использования его в качестве источника энергии только на начальном этапе работы мышцы в первую минуту, до момента активизации других более мощных источников. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Эта система определяет алактатную работоспособность мышц.
Максимальная алактатная мощность зависит от:
1. концентрации и активности фермента креатинфосфаткиназа (переносящего фосфатную группу с креатинфостфата на АДФ).
2. концентрации креатинфосфата.
Длительность удержания максимальной алактатной мощности составляет 6-12 секунд.
Алактатная емкость зависит от запасов креатинфосфата в мышце.
Эффективность креатинфосфаткиназной реакции очень велика (76%), так как реакция протекает непосредственно между двумя веществами на миофибриллах.
Лактатная (гликолитическая, лактацидная) система
Гликолиз – это процесс распада одной молекулы глюкозы на две молекулы молочной кислоты с выделением энергии, достаточной для фосфорилирования двух молекул АТФ, протекает в саркоплазме под воздействием 10 ферментов.
C6H12O6 + 2H3PO4 + 2АДФ → 2C3H6O3 + 2АТФ + 2H2O
Гликогенолиз – это процесс распада гликогена.
[C6H10O5] n + 3H3PO4 + 3АДФ → 2C3H6O3 + [C6H10O5] n-1 + 3АТФ + 2H2O
Для работы этой системы используются в основном внутримышечные запасы гликогена, а также глюкоза, поступающая из крови.
Гликолиз протекает без потребления кислорода и способен быстро восстанавливать запасы АТФ в мышце. Достигает максимума через 30-40 секунд интенсивной работы.
Эта система определяет лактатную работоспособность мышц.
Максимальная лактатная мощность определяется главным образом концентрацией и активностью ключевых ферментов гликолиза, которые зависят от:
1. устойчивости ферментов гликолиза к повышению кислотности среды, которая ингибирует их активность.
2. устойчивости кислотно-щелочного равновесия внутренней среды мышц, в условиях усиленной выработки молочной кислоты.
Время удержания максимальной мощности данного метаболического процесса составляет 60-180 секунд.
Гликолитическая емкость определяется главным образом запасами гликогена в мышцах, гликоген печени для процессов гликолиза не обладает достаточной мобильностью.
Метаболическая эффективность гликолиза оценивается значениями КПД порядка 0,35-0,52. Это означает, что почти половина всей выделяемой энергии превращается в тепло и не может быть использована в работе.
Умеренный сдвиг pH в кислую сторону активирует работу ферментов дыхательного цикла в митохондриях и усиливает аэробное энергообразование.
Значительное накопление молочной кислоты, появление избыточного СО2, изменение рН и гипервентиляция лёгких, отражающие усиление гликолиза в мышцах, обнаруживаются при увеличении интенсивности нагрузки более 50% максимальной аэробной мощности. Этот уровень нагрузки обозначается, как порог анаэробного обмена (ПАНО). Чем раньше он будет достигнут, тем быстрее вступит в действие гликолиз, сопровождающийся накоплением молочной кислоты и последующим развитием утомления работающих мышц.
Величина ПАНО является важным показателем эффективности энергообразования в мышцах, роста степени тренированности, который широко используется при биохимическом контроле функционального состояния спортсмена. С ростом степени тренированности на выносливость ПАНО увеличивается, т.е. наступает при более интенсивной работе.
Миокиназная реакция
«Аварийный» путь ресинтеза АТФ:
АДФ + АДФ → АТФ + АМФ
Происходит в мышцах при значительном увеличении концентрации АДФ в саркоплазме. Такая ситуация возникает при выраженном мышечном утомлении, когда другие пути ресинтеза АТФ уже не справляются.
Эта реакция так же обратима и используется для поддержания постоянного уровня АТФ в мышцах.
Аэробная (кислородная, окислительная) система.
В обычных условиях аэробный механизм ресинтеза АТФ обеспечивает около 90% общего количества АТФ, ресинтезируемой в организме.
Окисление протекает в митохондриях под воздействием специальных ферментов и требует затрат кислорода, а соответственно и времени на его доставку. Такие процессы называются аэробными. Окисление происходит в несколько этапов, сначала идет гликолиз (см. выше), но образовавшиеся в ходе промежуточного этапа этой реакции две молекулы пирувата не преобразуются в молекулы молочной кислоты, а проникают в митохондрии, где окисляются в цикле лимонной кислоты до углекислого газа и воды, давая энергию для производства еще 36 молекул АТФ.
C6H12O6 + 6O2 + 38АДФ + 38H3PO4 → 6CO2 + 44H2О + 38АТФ
Итого распад глюкозы по аэробному пути дает энергию для восстановления 38 молекул АТФ. Т.е. окисление в 19 раз эффективнее гликолиза. Если во время гликолиза организм усваивает в виде АТФ лишь 3% энергии, заложенной в молекуле глюкозы, то при аэробном окислении этот показатель равен 55% (включая те самые 3%). К тому же аэробное окисление может использовать более энергоемкие субстраты, такие как жиры, которые дают в 2 раза больше энергии, чем то же количество углеводов.
Субстратами окисления являются любые органические вещества: белки, жиры, углеводы. Долевое участие будет зависеть от характера работы:
| Интенсивность нагрузки | Преимущественно используются |
|---|---|
| до 50% МПК (лёгкая работа) | жиры |
| 60-90% МПК (тяжёлая работа) | углеводы |
| близкая к МПК | практически только углеводы |
Эта система определяет аэробную работоспособность мышц.
Максимальная аэробная мощность зависит главным образом от:
1. плотности митохондрий в мышечных волокнах;
2. концентрации и активности окислительных ферментов;
3. скорости поступления кислорода вглубь волокна.
Объем кислорода доступного для окислительных реакций лимитируется:
1. состоянием кардио-респираторной системы;
2. капилляризация мышц;
3. концентрация миоглобина;
4. диаметр мышечного волокна (чем меньше диаметр волокна, тем лучше оно снабжается кислородом и тем выше его относительная аэробная мощность).
Показатель количества кислорода, усваиваемого единицей массы тела за единицу времени – МПК (максимальное потребление кислорода).
Скорость производства АТФ за счет окисления достигает максимальных значений на 2-3-й минуте работы, что связано с необходимостью развертывания множества процессов, обеспечивающих доставку кислорода к митохондриям. Время удержания максимальной аэробной мощности составляет примерно 6 минут, в дальнейшем аэробная мощность снижается по причине усталости всех активно работающих систем организма.
Аэробная ёмкость очень высокая, т.к. для окисления используются любые органические вещества.
Метаболическая эффективность этого механизма – около 50%. Она определяется по ПАНО: у нетренированных людей ПАНО наступает при потреблении кислорода примерно 50% от МПК, а у высокотренированных на выносливость – при 80-90% от МПК.
Общая характеристика аэробных физических нагрузок
В спортивный практике к упражнениям аэробного характера относят длительные физические упражнения, где относительный вклад аэробного процесса в затратах энергии превышает 70%.
К аэробным циклическим упражнениям (по Я.М.Коцу, 1986) относятся:
1. упражнения максимальной аэробной мощности (3-10 минут);
2. упражнения близкой к максимальной аэробной мощности (10-30 минут);
3. упражнения субмаксимальной аэробной мощности (30-80 минут);
4. упражнения средней аэробной мощности (80-120 минут);
5. упражнения малой аэробной мощности (более 120 минут).
Аэробный механизм является основным при таких видах спорта, как: бег на дистанции 5-25 км, велогонки, плавание на 800-1500 м, бег на коньках на 5-10 км и др.
Ёмкость аэробного механизма, которая в значительной степени определяется запасами гликогена в скелетных мышцах и печени, а также уровнем утилизации кислорода мышцами, существенно повышается уже в течение 1,5-2 месяцев тренировки на выносливость.
Мощность аэробного механизма, которая зависит от МПК и активности окислительных ферментов, также увеличивается в процессе адаптации к мышечной деятельности через 2-3 месяца тренировок.
Аэробная направленность физических нагрузок происходит, как правило, в зоне умеренной мощности. При этом упражнения выполняются при максимуме аэробного производства энергии.
Кислородный запрос может достигать 500-1500 л, кислородный долг не превышает 5 л (до 10%). Содержание молочной кислоты в крови составляет 0,6-0,8 г*л-1. В ходе работы она может извлекаться тканями и аэробно окисляться в них.
Вследствие усиленного использования запасов гликогена в печени, содержание глюкозы в крови становится ниже 0,8 г*л-1. В моче в значительном количестве появляются продукты распада белков. Отмечается потеря организмом воды и минеральных солей.
Основными упражнениями для развития аэробных процессов энергообеспечения будут физические нагрузки, относящиеся к зоне большой и умеренной мощности с интенсивностью работы на уровне ПАНО и 100% МПК.
Работа энергетических систем во время аэробной физической нагрузки
Для большей вариабельности рассмотрим несколько вариантов аэробной физической нагрузки.
При беге на длинные дистанции (5 и 10 км) аэробное окисление углеводов является основным механизмом энергообеспечения работы, так как на его долю приходится до 87% общих затрат энергии на дистанции 5 км и 97% на дистанции 10 км.
Вклад анаэробных источников на этих дистанциях также достаточно большой. Он может достигать 15% общих затрат энергии и играет важную роль при финишном ускорении, приносящем победу при беге на длинные дистанции.
Наиболее значительным фактором, влияющим на выносливость, является кислородное снабжение работающих мышц, поскольку потребление кислорода во время бега поддерживает максимальную скорость окисления углеводов. Порог анаэробного обмена у стайеров при работе достигается при 75-90% МПК.
При марафонском беге затраты энергии восполняются исключительно за счёт аэробного процесса. Погашение этих затрат невозможно только за счёт окисления углеводов из-за недостаточности запасов гликогена в работающих мышцах спортсмена, поэтому значительная часть энергии образуется за счёт окисления жиров, на долю которых может приходится от 10 до 50% общих затрат энергии.
Вклад жиров на длинных и сверхдлинных дистанциях у высокотренированных бегунов с большими запасами гликогена в работающих мышцах составляет 12-20%, у нетренированных бегунов – более 80%. Всего на дистанции марафонского бега окисляется около 300 г жиров.
Использование жиров в качестве источника энергии менее эффективно по сравнению с окислением углеводов, так как происходит с более низкой скоростью и с большим потреблением кислорода.
Рис 2. Механизмы энергообеспечения бега на 10 000 м и марафонского бега (пунктирная черта показывает момент исчерпания запасов гликогена)
При длительной работе наряду с увеличением использования в энергетическом обмене жиров может происходить новообразование углеводов из веществ неуглеводной природы (глюконеогенез).
Основным субстратом глюконеогенеза являются аминокислоты, часть которых накапливается в мышце при работе в результате распада тканевых белков.
Рассмотрим, как включаются в работу энергетические системы во время аэробных физических нагрузок.
Таблица 2. Вклад различных источников энергии в обеспечение ресинтеза АТФ при беге в аэробном режиме работы (в %).
| Дистанция, м | Креатинфосфат | Анаэробное окисление гликогена мышц | Аэробное окисление гликогена мышц | Глюкоза крови (гликоген печени) | Жирные кислоты |
|---|---|---|---|---|---|
| 1500 | Минимальный | 25 | 75 | — | — |
| 5000 | Минимальный | 12,5 | 87,5 | — | — |
| 10000 | Минимальный | 3 | 97 | — | — |
| Марафон | — | — | 75 | 5 | 20 |
| Супермарафон (84 км) | — | — | 35 | 5 | 60 |
| 24-часовой забег | — | — | 10 | 2 | 88 |
При переходе из состояния покоя к мышечной деятельности потребности в кислороде возрастает в несколько раз, но сразу она не может быть удовлетворена. Необходимо время, чтобы усилилась деятельность кардиореспираторной системы, чтобы кровь, обогащённая кислородом смогла дойти до работающих мышц. По мере усиления активности работы этих систем постепенно увеличивается потребление кислорода в работающих мышцах. Скорость потребления кислорода увеличивается до тех пор, пока не наступит истинное устойчивое состояние метаболических процессов, при котором потребление кислорода в данный момент времени точно соответствует потребности организма в нём (кислородному запросу).
До этого момента потребность организма в энергии обеспечивается большей частью за счёт работы анаэробных энергетических систем. Как мы уже отмечали выше, скорость развёртывания креатинфосфатной системы до полной мощности – доли секунды, лактатной – около половины минуты. В зависимости от того, какой кислородный запрос работы имеет нагрузка, кислородный дефицит на начальном её этапе восполняется за счёт разного участия анаэробных систем, но в любом случае развёртывания этих систем на полную мощность при нагрузках аэробного характера не требуется. В результате происходит накопление в организме недоокисленных продуктов анаэробного распада.
Рис 3. Кислородный приход, кислородный дефицит и кислородный долг при аэробной работе (а) лёгкой, (б) тяжёлой интенсивности. 1 – быстрый, 2 – медленный компоненты кислородного долга.
При работе в устойчивом состоянии часть анаэробных метаболитов может окисляться за счёт усиления аэробных реакций в процессе работы, а другая их часть устраняется после работы.
При выполнении работы с уровнем запроса около 50% МПК прирост концентрации молочной кислоты невелик (до 0,4-0,5 г/л), а при выполнении продолжительных нагрузок с уровнем запроса 50-85% МПК, возрастает до 1-1,5 г/л. Концентрация молочной кислоты значительно возрастает в первые 2-10 минут работы, а затем либо остаётся на прежнем уровне, либо снижается. То есть максимальная концентрация молочной кислоты в крови наблюдается до тех пор, пока не установилось устойчивое состояние, создающее условия для аэробного её окисления.
Для восстановления энергетических источников и окисления недоокисленных продуктов требуется дополнительное количество кислорода, поэтому некоторое время после окончания работы потребление его продолжает оставаться повышенным по сравнению с уровнем покоя. Этот излишек потребления кислорода в период восстановления получил название «кислородный долг».
Кислородный долг всегда больше кислородного дефицита. Чем больше интенсивность и продолжительность работы, тем кислородный долг выше.
После работы в устойчивом состоянии кислородный долг наполовину восполняется уже за 30 секунд, а полностью через 3-5 минут. После интенсивной работы «погашение» долга происходит в две фазы.
Быстрый (алактатный) компонент кислородного долга включает то количество кислорода, которое необходимо для ресинтеза АТФ и креатинфосфота. Он характеризует вклад креатинфосфатной энергетической системы в обеспечении выполненной работы.
Медленный (лактатный) компонент кислородного долга включает то количество кислорода, которое необходимо для окисления образовавшейся молочной кислоты при выполнении работы. Его величина характеризует участие лактатной энергетической системы, а при длительной работе – и других процессов, долю которых оценить весьма затруднительно. Медленный компонент устраняется наполовину за 15-25 минут, а полностью – за 1,5-2 часа.
Подводя итог, хочется отметить следующее:
• во время аэробной физической нагрузки работают все энергетические системы организма, но подавляющую роль играет аэробная система;
• все системы начинают работать одновременно с началом нагрузки, но за счёт разной скорости развёртывания процессов энергообразования, аэробная система полностью обеспечивает кислородный запрос не сразу, и на начальном этапе (несколько минут) кислородный приход компенсируют анаэробные энергетические системы.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности – Киев: Олимпийская литература, 2000
2. Граевская Н.Д., Долматова Т.И. Спортивная медицина: Курс лекций и практические занятия. В 2х частях. – М.: Советский спорт, 2004
3. Ким Н.К., Дьяконов М.Б. Фитнес. Учебник – М.: Советский спорт, 2006
4. Макарова Г.А. Медицинский справочник тренера – М.: Советский спорт, 2004
5. Руненко С.Д. Фитнес: мифы, иллюзии, реальность – М.: Советский спорт, 2005
6. Протасенко В.А. Думай! Или ‘Супертренинг’ без заблуждений, — журнал «Мускуляр», 2001
7. Хоули Э., Дон Френкс Б. Руководство инструктора оздоровительного фитнеса – Киев. Олимпийская литература, 2004
(С)2011 Д.А.Жабкин
Содержание
Энергетические субстратыПравить
Работа мышц требует постоянного поступления АТФ для поддержания цикла формирования поперечных связей. При этом миозин выступает в роли АТФазы и расщепляет АТФ на АДФ и неорганический фосфат (Ф). Мышцы имеют очень ограниченные резервы АТФ (этого запаса может хватить только на 4-6 с сократительной активности), но могут регенерировать АТФ за очень короткое время за счет присоединения фосфатной группы к АДФ.
Процесс быстрой регенерации АТФ в мышцах происходит при переносе фосфатной группы с креатинфосфата на АДФ с образованием АТФ и креатинина (такой способ ресинтеза АТФ называют анаэробно-алактатным энергообеспечением). Поскольку наличный запас креатинфосфата в мышечной клетке невелик (его хватает на 6-10 с интенсивной работы), для более длительного функционирования мышц необходим синтез нового АТФ в ходе анаэробного гликолиза (анаэробно-гликолитическое энергообеспечение), при котором 1 моль глюкозы расходуется на синтез 2 молей АТФ, либо в ходе окислительного фосфорилирования в митохондриях (аэробное энергообеспечение), при котором за счет окисления 1 моля глюкозы синтезируется 34 моля АТФ. Наряду с глюкозой в качестве источника энергии мышцы могут использовать триглицериды в процессе бета-окисления жирных кислот.
Регенерация аденозинтрифосфата из креатинфосфатаПравить
Резервов креатинфосфата в мышцах достаточно для регенерации АТФ в течение не более 10 с. Продукт метаболизма креатина — креатинин — затем выводится в кровь и попадает в мочу. Суточная продукция креатинина зависит от мышечной массы, поэтому уровень креатинина в плазме у мужчин выше, чем у женщин. Плазменный уровень креатинина также сильно зависит от общей тренированности. В норме у нетренированных лиц он составляет 0,5-1,2 мг/дл для мужчин и 0,5-1 мг/дл для женщин. Из организма креатинин выводится почками, повышение его концентрации в плазме крови может говорить о нарушении функции почек, однако значительное повышение уровня креатинина наблюдают только при тяжелой почечной патологии.
Запомните: АТФ — энергетический субстрат мышечной ткани. Резервов АТФ в мышцах хватает только на 5-6 с, за счет резервов креатинфосфата регенерация АТФ возможна еще 10-20 с (анаэробно-алактатное энергообеспечение). АТФ синтезируется в процессе гликолиза (анаэробно-гликолитическое энергообеспечение), а также окислительного фосфорилирования (аэробное энергообеспечение).
Анаэробный синтез аденозинтрифосфата в процессе гликолизаПравить
При отсутствии или недостатке кислорода (анаэробные условия) мышца может регенерировать АТФ за счет процесса гликолиза. Такие условия возникают, как правило, в начале циклической мышечной работы (врабатывание), а также в том случае, если величина физической нагрузки больше, чем скорость образования энергии за счет аэробного энергетического процесса. При этом из глюкозы в цитоплазме мышечной клетки образуется метаболит пируват, а конечным продуктом является молочная кислота. Мышцы получают глюкозу из крови или за счет распада мышечного гликогена. Молекула глюкозы представляет собой 6-атомный спирт (т. е. каркас молекулы состоит из 6 атомов углерода). В процессе перегруппировки атомов и расщепления молекула глюкозы распадается на 2 молекулы пировиноградной кислоты (пирувата), каждая из которых содержит по 3 атома углерода. Пируват представляет собой один из типичных субстратов для работы митохондрий, но если он не успевает туда проникнуть или в клетке наблюдается нехватка кислорода, то в этом случае из пирувата в цитоплазме клетки образуется молочная кислота, легко распадающаяся на анион лактата- и Н+. Выходящий в кровь по градиенту концентрации лактат обусловливает локальное закисление за счет повышения концентрации катионов водорода. В результате этих биохимических превращений из 1 моля глюкозы образуется 2 моля АТФ.
Мышечные волокна способны накапливать глюкозу в виде гликогена. Гликоген представляет собой сильно разветвленную молекулу, что обеспечивает быстрый доступ расщепляющих ферментов к фрагментам молекулы гликогена и быстрое высвобождение запасов глюкозы. Однако для длительной мышечной активности (> 20 мин) мышцы должны получать АТФ в ходе окисления липидов, т. к. запасы мышечного гликогена истощаются. Именно по этой причине считается, что для профилактики и борьбы с ожирением нужны длительные циклические нагрузки аэробного характера.
Аэробный синтез аденозинтрифосфата в процессе окислительного фосфорилированияПравить
Пути энергообеспечения мышечной ткани
При наличии кислорода пируват подвергается окислительному фосфорилированию в цикле трикарбоновых кислот до С02 и Н20. При этой реакции возможно синтезировать больше АТФ, чем при гликолизе — из 1 моля глюкозы образуется 34 моля АТФ, — однако этот процесс более медленный: скорость образования АТФ в аэробном процессе почти в 2 раза ниже, чем в процессе анаэробного гликолиза.
Важность наличия различных систем синтеза АТФ определяется различным временем регенерации АТФ (рис.). Так, на коротких дистанциях (60, 100 м) особую роль играет креатинфосфат, процессы анаэробного гликолиза достигают максимума через 30 с — 1 мин после начала нагрузки и сохраняют активность к концу средних дистанций (200, 400 м), а на длинных дистанциях наибольшее поступление АТФ обеспечивает окислительное фосфорилирование, которое достигает максимума через 1-3 мин. На время регенерации АТФ также оказывают влияние общая тренированность и питание.
Синтез аденозинтрифосфата в процессе β-окисления жирных кислотПравить
Другим источником получения энергии в мышцах является β-окисление жирных кислот. Свободные жирные кислоты поступают в мышцы из крови и накапливаются в них в виде триглицеридов. Триглицериды являются эфирами глицерина и трех жирных кислот различной длины.
По сравнению с очень малыми запасами гликогена в организме (около 500 г) запасы жира составляют 12 кг, однако выход АТФ из гликогена практически в 2 раза превышает эффективность окисления жирных кислот. Таким образом, последний вариант получения АТФ «выгоден», когда можно удовлетворить большие потребности в энергии и сохранить при этом «ценные» запасы гликогена. Сэкономленные запасы гликогена могут использоваться при дополнительном краткосрочном повышении нагрузки, например при промежуточном или конечном спурте при беге.
В ходе окисления жирных кислот для разрушения ненасыщенных двойных связей в цепи жирных кислот необходимо больше кислорода, однако это играет второстепенную роль в энергетическом балансе организма.
Энергетический обмен в мышцахПравить
Факторы, влияющие на энергетический обмен
С точки зрения физики энергия имеет такую же размерность, как работа, а работа — это сила, умноженная на расстояние. Количество работы, выполняемой за единицу времени, называют мощностью. Энергетический обмен (преобразование энергии в организме) — это мощность термодинамических процессов, поскольку он выражает выполненную работу (или потраченную энергию) за единицу времени. Основными проявлениями энергетического обмена в организме являются мышечная работа и выделяющееся тепло. Организм человека подчиняется закону сохранения энергии, в соответствии с которым количество поглощенной энергии (в различных формах) и образовавшейся энергии (главным образом в виде тепла и механической работы) равны. Сбалансированный процесс поглощения и отдачи энергии организмом называют энергетическим обменом. Измерение энергетического обмена играет большую роль для оценки объема физической нагрузки (например, при изучении физиологии спорта и труда). На энергетический обмен влияет множество факторов (рис.).
Энергетический обмен при физической нагрузкеПравить
Энергетический обмен даже при минимальной физической нагрузке становится в 1,5-2 раза выше, чем в условиях покоя. Чем сильнее повышается энергетический обмен при выполнении физической нагрузки, тем короче то время, которое человек способен такую нагрузку выполнять. Максимальное увеличение скорости обменных процессов у человека может быть примерно 30-кратным (по сравнению с основным обменом), но время удержания такой нагрузки не превышает 6 с. Если скорость метаболических процессов увеличена в 10 раз по сравнению с основным обменом, то такую циклическую нагрузку нетренированный человек способен удерживать 3-4 мин, а спортсмен, тренированный на выносливость, — до 6 мин. Считается, что именно в таком режиме реализуются максимальные аэробные возможности человека.
Запомните: Энергетический обмен (основной обмен, обмен при физической нагрузке) определяют с помощью непрямой калориметрии по объему потребления кислорода.
Читайте такжеПравить
- Энергетические процессы в мышце
- Энергетический баланс
- Энергообеспечение мышечной деятельности
- Обмен покоя
- Основной обмен
- Коэффициент полезного действия мышц
При
выполнении упражнений преимущественно
аэробного характера скорость
потребления кислорода (л О2/мин)
тем выше, чем больше мощность выполняемой
нагрузки (скорость перемещения). Поэтому
в видах спорта, требующих проявления
большой выносливости, спортсмены должны
обладать большими аэробными возможностями:
1)
высокой максимальной скоростью
потребления кислорода, т.
е. большой аэробной «мощностью», и
2)
способностью длительно поддерживать
высокую скорость потребления
кислорода (большой
аэробной «емкостью»).
Максимальное
потребление кислорода МПК.
Аэробные
возможности человека определяются прежде
всего максимальной для него скоростью
потребления кислорода. Чем выше МПК,
тем больше абсолютная мощность
максимальной аэробной нагрузки. Кроме
того, чем выше МПК, тем относительно
легче и потому длительнее выполнение
аэробной работы
Таким
образом, чем выше МПК у спортсмена, тем
более высокую скорость он может
поддерживать на дистанции, тем,
следовательно, выше (при прочих равных
условиях) его спортивный результат в
упражнениях, требующих проявления
выносливости. Чем выше МПК, тем больше
аэробная работоспособность (выносливость),
т. е. тем больший объем работы аэробного
Характера способен выполнить человек.
Причем эта зависимость выносливости
от МПК проявляется (в некоторых пределах)
тем больше, чем меньше относительная
мощность аэробной нагрузки.
Абсолютные
показатели МПК (л
О2/мин)
находятся в прямой связи с размерами
(весом) тела. Поэтому наиболее высокие
абсолютные показатели МПК имеют гребцы,
пловцы, велосипедисты, конькобежцы. В
этих видах спорта наибольшее значение
для физиологической оценки данного
качества имеют абсолютные показатели
МПК.
Относительные
показатели МПК (мл
О2/кг
* мин) у высококвалифицированных
спортсменов находятся в обратной
зависимости от веса тела. При беге и
ходьбе выполняется значительная работа
по вертикальному перемещению массы
тела и, следовательно, при прочих равных
условиях (одинаковой скорости передвижения)
чем больше вес спортсмена, тем больше
совершаемая им работа (потребление О2).
Поэтому бегуны на длинные дистанции,
как правило, имеют относительно небольшой
вес тела (прежде всего за счет минимального
количества жировой ткани и относительно
небольшого веса костного скелета).
Уровень
МПК зависит от максимальных возможностей
двух функциональных систем:
1)
кислородтранспортной системы, абсорбирующей
кислород из окружающего воздуха и
транспортирующей его к работающим
мышцам и другим активным органам и
тканям тела;
2)
системы утилизации кислорода, т.
е. мышечной системы, экстрагирующей и
утилизирующей доставляемый кровью
кислород. У спортсменов, имеющих высокие
показатели МПК, обе эти системы обладают
большими функциональными возможностями.
Упражнения
максимальной аэробной мощности (с
дистанционным потреблением кислорода
95-100% от индивидуального МПК) -это
упражнения, в которых преобладает
аэробный компонент энергопродукции —
он составляет до 60-70%. Однако энергетический
вклад анаэробных (преимущественно
гликолитических) процессов еще очень
значителен. Основным энергетическим
субстратом при выполнении этих упражнений
служит мышечный гликоген, который
расщепляется как аэробным, так и
анаэробным путем (в последнем случае с
образованием большого количества
молочной кислоты). Предельная
продолжительность таких упражнений —
3-10 мин. К соревновательным упражнениям
этой группы относятся: бег на 1500 и 3000 м,
бег на 3000 и 5000 м на коньках, плавание на
400 и 800 м, академическая гребля (классические
дистанции), заезды на 4 км на велотреке.
Через 1,5-2 мин после начала упражнений
достигаются максимальные для данного
человека ЧСС, систолический объем крови
и сердечный выброс, скорость потребления
кислорода (МПК). После окончания упражнения
концентрация лактата в крови Достигает
15-25 ммоль/л в обратной зависимости от
предельной продолжительности упражнения
и в прямой — от квалификации-спортсмена
(спортивного результата). Ведущие
физиологические системы имеханизмы —
общие для всех аэробных упражнений;
кроме того, существенную роль играет
мощность лактацидной (гликолитической)
энергетической системы рабочих мышц.
36.
Нагрузка и отдых при занятиях физическими
упражнениями. Дозирование физической
нагрузки.
Нагрузка —
определяющая величина воздействия
физических упражнений на организм
человека.
Нагрузку
рассматривают с двух сторон:
-
Внешняя —
определяется показателями нагрузки,
характерными внешними параметрами
совершаемой работы, т.е.:-
Продолжительностью;
-
Объемом
выполнения; -
Количеством
повторений.
-
-
Внутренняя —
определяется величиной физиологических
и биохимических сдвигов в организме,
вызываемых упр-нием, т.е.:-
Степенью
увеличения ЧСС; -
Минутным
объем крови; -
Потреблением
кислорода.
-
Нагрузка
бывает:
-
Стандартной –
одинаковой по своим внешним параметрам
в каждый момент выполнения упражнения. -
Вариативной (переменной)
– изменяющейся в ходе выполнения
упражнения.
Величина
нагрузки складывается из объема и
интенсивности выполнения упражнений
за одну тренировку или тренировочный
цикл.
Под объемом понимают.
-
Длительность
выполнения упр-ний. -
Суммарное
количество выполняемой работы (за
одно занятие или месяц).
Например:
-
в
упражнениях циклического характера
объем оценивают по километражу; -
в
упражнениях с отягощениями по количеству
подходов; -
в
спортивных играх по количеству атакующих
и защитных действий.
интенсивность —
это скорость выполнения определенного
объема нагрузки.
Чем
выше интенсивность упражнения, тем
меньше возможный объем нагрузки.
Например:
-
объем
передвижения (в
беге, плавание); -
темп
игры (спортивные
игры).
Моторная
плотность –
отношение времени, потраченного на
выполнение упражнения к общему времени
занятия.
Нагрузка
имеет характер:
-
Непрерывный —
когда при выполнении упражнения
отсутствует пауза отдыха. -
Прерывный (интервальный)
— когда между упражнениями имеются
интервалы отдыха.
Отдых может
быть:
-
Пассивный —
относительный покой и отсутствие
активной деятельности. -
Активный —
переключение на другой вид деятельности
(в беге — спокойная ходьба; в плавание
– неторопливые движения в воде). -
Смешанный –
сочетание активного и пассивного
отдыха.
Интервалы
отдыха:
-
Полный —
обеспечивает восстановление
работоспособности до исходного уровня,
т.е. появляется чувство готовности к
работе. -
Неполный —
очередное выполнение упражнения
приходится на период не довосстановления
работоспособности. -
Экстремальный —
очередное выполнение упражнения
совпадает с фазой повышения
работоспособности. -
Длинный —
очередная работа выполнена в тот момент,
когда следы предыдущего задания почти
утрачены.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Дано определение ресинтеза АТФ. Описаны основные пути ресинтеза АТФ в мышечных волокнах: креатинфосфатный, гликолитический, миокиназный и тканевое дыхание. Описаны количественные критерии путей ресинтеза АТФ, соотношение между различными путями ресинтеза АТФ при мышечной работе, а также между путями ресинтеза АТФ и зонами относительной мощности.
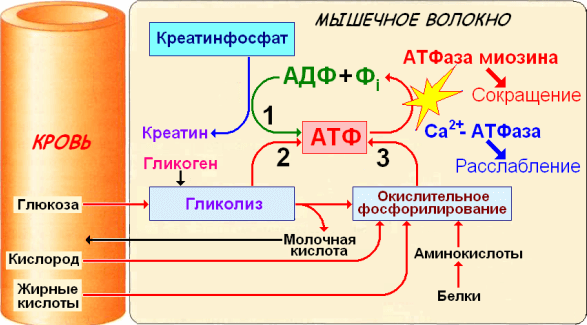
Ресинтез АТФ в мышечных волокнах
Определение
Ресинтез АТФ – синтез АТФ из различных энергетических субстратов во время физической работы в мышечных волокнах.
Формула ресинтеза АТФ выглядит следующим образом:
АДФ+фосфат+энергия → АТФ.
Пути ресинтеза АТФ
Ресинтез АТФ может осуществляться двумя путями:
- с участием кислорода (аэробный путь).
- без участия кислорода (анаэробный путь);
Аэробный путь (тканевое дыхание, аэробное или окислительное фосфорилирование) – основной способ образования АТФ в мышечных волокнах. Он протекает в митохондриях мышечных волокон. В результате тканевого дыхания выделяется 39 молекул АТФ. Окисляемое вещество распадается до углекислого газа и воды.
Анаэробный ресинтез АТФ
Анаэробные пути ресинтеза АТФ являются дополнительными способами образования АТФ в мышечных волокнах в тех случаях, когда основной путь получения АТФ – тканевое дыхание не может обеспечить мышечную деятельность необходимым количество кислорода. Эти механизмы ресинтеза АТФ активно функционируют в начале выполнения физических упражнений, когда тканевое дыхание не полностью «развернулось», а также при физических нагрузках высокой мощности.
Анаэробный ресинтез АТФ в мышечных волокнах возможен посредством нескольких механизмов:
- Креатинфосфатный ресинтез АТФ – ресинтез АТФ из креатинфосфата;
- Гликолитический ресинтез АТФ – ресинтез АТФ из гликогена мышц;
- Миокиназный (аденилаткиназный) ресинтез АТФ – ресинтез АТФ из АДФ при значительном накоплении в мышечных волокнах АДФ. Рассматривается как аварийный механизм, обеспечивающий ресинтез АТФ, когда другие пути ресинтеза АТФ невозможны.
Количественные критерии путей ресинтеза АТФ
Существуют количественные критерии путей ресинтеза АТФ. К ним можно отнести: максимальную мощность, время развертывания, время сохранения или поддержания максимальной мощности, метаболическую ёмкость (табл. 1).
- Максимальная мощность – максимальное количество АТФ, которое может образоваться в единицу времени при функционировании данного пути ресинтеза АТФ.
- Время развертывания – минимальная длительность, необходимая для выхода ресинтеза АТФ на свою максимальную мощность.
- Время сохранения или поддержания максимальной скорости – длительность функционирования данного пути ресинтеза АТФ с максимальной мощностью.
- Метаболическая ёмкость – количество АТФ, которое может образоваться во время мышечной работы за счёт данного пути ресинтеза АТФ.
Таблица 1. Количественные критерии основных путей ресинтеза АТФ (С.С. Михайлов, 2009)
| Пути ресинтеза АТФ | Максимальная мощность, кал/мин кг | Время развертывания | Время сохранения максимальной мощности | Метаболическая ёмкость |
| Креатинфосфатный | 900-1100 | 1-2 с | 8-10 с | |
| Гликолитический | 750-850 | 20-30 с | 2-3 мин. | При анаэробном окислении гликогена образуются 3 молекулы АТФ в расчете на одну молекулу глюкозы |
| Аэробный | 350-450 | 3-4 мин. | Десятки минут | При аэробном окислении гликогена образуются 39 молекул АТФ в расчете на одну молекулу глюкозы (самый экономичный) |
Соотношение между различными путями ресинтеза АТФ
При любой мышечной работе функционируют все три основных механизма ресинтеза АТФ, но включаются они последовательно. В первые секунды ресинтез АТФ осуществляется за счет креатинфосфатной реакции, затем включается гликолиз. По мере продолжения работы на смену гликолизу приходит тканевое дыхание (рис.1). Эта смена механизмов ресинтеза АТФ приводит к уменьшению суммарной выработки АТФ.
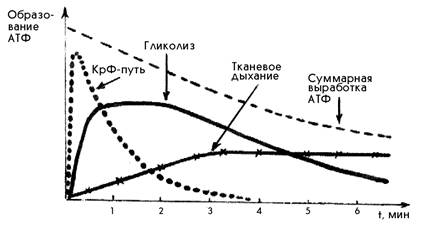
Пути ресинтеза АТФ и зоны относительной мощности
В.С. Фарфель приводит следующее соотношение мощности работы и основной системы энергообеспечения (табл.2)
Таблица 2. Зоны мощности работы и основная система энергообеспечения (В.С. Фарфель)
| Мощность работы | Основная система энергообеспечения | Типичное время работы |
| Максимальная | Креатинфосфатная реакция | до 20 с |
| Субмаксимальная | Гликолиз | до 5 мин. |
| Большая | Гликолиз+ тканевое дыхание | до 30 мин. |
| Умеренная | Тканевое дыхание | Более 30 мин. |
J.T. Cramer (2008) приводит несколько иное соотношение зон мощности и основных систем энергообеспечения (табл.3)
Таблица 3. — Зоны относительной мощности и основная ситема энергообеспечения (J.T. Cramer, 2008)
| % от максимальной мощности работы | Основная система энергообеспечения | Время работы |
| 90-100 | Креатинфосфатная реакция | 5-10 с |
| 75-90 | Гликолиз | 15-30 с |
| 30-75 | Гликолиз+ тканевое дыхание | 1-3 мин. |
| 20-30 | Тканевое дыхание | Более 3 мин. |
Видео про гидролиз и ресинтез АТФ
Литература
- Михайлов С.С. Спортивная биохимия. – М.: Советский спорт, 2009.– 348 с.
- Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности.- Киев: Олимпийская литература, 2000.- 504 с.
С уважением, А.В.Самсонова
Похожие записи:
Сила тяжести
Дано определение силы тяжести. Показано, что сила тяжести является частным случаем силы гравитации. Описаны факторы, определяющие силу тяжести:…
Сила
Дано определение силы в механике. Описаны факторы, определяющие действие на тело силы: направление, точка приложения и численное значение.
Звенья тела человека как рычаги
Дано описание опорно-двигательного аппарата (ОДА) человека как системы рычагов. Приведен пример расчета силы двуглавой мышцы плеча…
Метаболический стресс. Накопление лактата в мышцах
Описан механизм влияния метаболического стресса (накопления лактата) на гипертрофию мышечных волокон. Показано, что накопление лактата приводит к…
Механическое повреждение мышечных волокон
Описаны механизмы механического повреждения мышечных волокон при силовой тренировке, приводящие к гипертрофии скелетных мышц. Показано, что…
Механическое напряжение (механотрансдукция) в скелетных мышцах
Описаны процессы передачи механического напряжения в скелетных мышцах. Показано, что механическое напряжение, возникающее вследствие сокращения скелетных…
|
Материалы старого сайта находятся здесь |
 
|
Энергетика мышечной деятельности — 1
Автор: —
Дата: 2010-03-28
Ни одно движение не может быть выполнено без затрат энергии. Единственным универсальным и прямым источником энергии для мышечного сокращения служит аденозинтрифосфат -АТФ; без него поперечные «мостики лишены энергии и актиновые нити не могут скользить вдоль миозиновых, сокращения мышечного волокна не происходит. АТФ относится к высокоэнергетическим (макроэргическим) фосфатным соединениям, при расщеплении (гидролизе) которого выделяется около 10 ккал/кг свободной энергии. При активизации мышцы происходит усиленный гидролиз АТФ, поэтому интенсивность энергетического обмена возрастает в 100-1000 раз по сравнению с уровнем покоя. Однако, запасы АТФ в мышцах сравнительно ничтожны и их может хватить лишь на 2-3 секунды интенсивной работы. В реальных условиях для того, чтобы мышцы могли длительно поддерживать свою сократительную способность, должно происходить постоянное восстановление (ресинтез) АТФ с той же скоростью, с какой он расходуется. В качестве источников энергии при этом используются углеводы, жиры и белки. При полном или частичном расщеплении этих веществ освобождается часть энергии, аккумулированная в их химических связях. Эта освободившаяся энергия и обеспечивает ресинтез АТФ (см. табл.).
Энергетические резервы человека ( с массой тела 75 кг)
| Источники энергии | Энергоемкость, кДж | Возможная продолжительность работы, с |
| АТФ | 4 — 5 | 2 — 3 |
| Креатинфосфат (КрФ) | 14 — 15 | 15 — 20 |
| Гликоген+глюкоза | 4600 — 13000 | 120 — 240 |
| Жиры | 300000 — 400000 | более 240 |
Биоэнергетические возможности организма являются наиболее важным фактором, лимитирующим его физическую работоспособность. Образование энергии для обеспечения мышечной работы может осуществляться анаэробным (бескислородным) и аэробным (окислительным) путем. В зависимости от биохимических особенностей протекающих при этом процессов принято выделять три обобщенных энергетических системы, обеспечивающих физическую работоспособность человека:
алактная анаэробная, или фосфагенная, связанная с процессами ресинтеза АТФ преимущественно за счет энергии другого высокоэнергетического фосфатного соединения — креатинфосфата (КрФ);
гликолитическая (лактацидная) анаэробная, обеспечивающая ресинтез АТФ и КрФ за счет реакций анаэробного расщепления гликогена или глюкозы до молочной кислоты (МК);
аэробная (окислительная), связанная с возможностью выполнения работы за счет окисления энергетических субстратов, в качестве которых могут использоваться углеводы, жиры, белки при одновременном увеличении доставки и утилизации кислорода в работающих мышцах.
Каждый из перечисленных биоэнергетических компонентов физической работоспособности характеризуется критериями мощности, емкости и эффективности (см. рис.1).

Рис. 1. Динамика скорости энергопоставляющих процессов в работающих мышцах в зависимости от продолжительности упражнения (по Волковоу Н.И., 1986)
Критерий мощности оценивает то максимальное количество энергии в единицу времени, которое может быть обеспечено каждой из метаболических систем.
Критерий емкости оценивает доступные для использования общие запасы энергетических веществ в организме, или общее количество выполненной работы за счет данного компонента.
Критерий эффективности показывает, какое количество внешней (механической) работы может быть выполнено на каждую единицу затрачиваемой энергии.
Фосфагенная система представляет собой наиболее быстро мобилизуемый источник энергии. Ресинтез АТФ за счет креатинфосфата во время мышечной работы осуществляется почти мгновенно. При отщеплении фосфатной группы от КрФ высвобождается большое количество энергии, которая непосредственно используется для восстановления АТФ. Поэтому КрФ является самым первым энергетическим резервом мышц, используемым как немедленный источник регенерации АТФ. АТФ и КрФ действуют как единая система энергоснабжения мышечной деятельности. Эта система обладает наибольшей мощностью по сравнению с гликолитической и аэробной, и играет основную роль в обеспечении кратковременной работы предельной мощности, осуществляемой с максимальными по силе и скорости сокращениями мышц: при выполнении кратковременных усилий «взрывного» характера, спуртов, рывков, как, например, спринтерский бег, прыжки, метания или удары рукой и ногой в рукопашном бою и т. п. Наибольшая мощность алактатного анаэробного процесса достигается в упражнениях продолжительностью 5-6 секунд и у высоко подготовленных спортсменов достигает уровня 3700 кДж/кГ в минуту. Однако емкость этой системы невелика в связи с ограниченностью запасов АТФ и КрФ в мышцах. Вместе с тем, время удержания максимальной анаэробной мощности зависит не столько от емкости фосфагенной системы, сколько от той ее части, которая может быть мобилизована при работе с максимальной мощностью. Расходуемое количество КрФ во время выполнения упражнений максимальной мощности составляет всего лишь примерно одну треть от его общих внутримышечных запасов. Поэтому продолжительность работы максимальной мощности обычно даже у высококвалифицированных спортсменов не превышает 15-20 секунд.
Анаэробный гликолиз начинается практически с самого начала работы, но достигает своей максимальной мощности лишь через 15-20 секунд работы предельной интенсивности, и эта мощность не может поддерживаться более 2.5 — 3.0 минут.
Гликолитическая анаэробная система характеризуется достаточно большой мощностью, достигая у высокотренированных людей уровня примерно 2500 кДж/кГ в минуту. Энергетическими субстратами при этом служат углеводы — гликоген и глюкоза. Гликоген, запасаемый в мышечных клетках и печени — это цепочка молекул глюкозы (глюкозных единиц). При расщеплении гликогена его глюкозные единицы последовательно отщепляются. Каждая глюкозная единица из гликогена восстанавливает 3 молекулы АТФ, а молекула глюкозы — только 2 молекулы АТФ. Из каждой молекулы глюкозы образуется 2 молекулы молочной кислоты (МК). Поэтому при большой мощности и продолжительности гликолитической анаэробной работы в мышцах образуется значительное количество МК. Накапливающаяся в работающих мышечных клетках МК легко диффундирует в кровь и, до определенной степени концентрации, связывается буферными системами крови для сохранения внутренней среды организма (гомеостазиса). Если количество МК, образующейся в процессе выполнения работы гликолитической анаэробной направленности, превышает возможности буферных систем крови, то это приводит к их быстрому исчерпанию и вызывает сдвиг кислотно-щелочного равновесия крови в кислую сторону. В конечном итоге, это вызывает угнетение ключевых ферментов анаэробного гликолиза, вплоть до полного торможения их активности. При этом снижается скорость и самого гликолиза. Значительное закисление приводит также к уменьшению скорости алактатного анаэробного процесса и общему снижению мощности работы.
Продолжительность работы в гликолитическом анаэробном рижиме лимтируется в основном не количеством (емкостью) ее энергетических субстратов, а уровнем концентрации МК и степенью тканевой адаптации к кислотным сдвигам в мышцах и крови. Во время выполнения мышечной работы, обеспечиваемой преимущественно анаэробным гликолизом, резкого истощения мышечного гликогена и глюкозы в крови и печени не происходит. В процессе физической подготовки гипогликемия (снижение концентрации глюкозы в крови) может возникнуть по другим причинам.Для высокого уровня проявления гликолитической анаэробной способности (специальной выносливости) существенное значение имеет степень тканевой адаптации к происходящим при этом сдвигам кислотно-щелочного равновесия. Здесь особо выделяется фактор психической устойчивости, который позволяет при напряженной мышечной деятельности волевым усилием преодолевать возникающие с развитием утомления болезненные ощущения в работающих мышцах и продолжать выполнять работу, несмотря на усиливающееся стремление к ее прекращению.
При переходе от состояния покоя к мышечной деятельности потребность в кислороде (его запрос) возрастает во много раз. Однако, необходимо по крайней мере 1-2 минуты, чтобы усилилась деятельность кардио-респираторной системы, и обогащенная кислородом кровь могла быть доставлена к работающим мышцам. Потребление кислорода работающими мышцами увеличивается постепенно, по мере усиления деятельности систем вегетативного обеспечения. С увеличением длительности упражнения до 5-6 минут быстро наращивается скорость процессов аэробного образования энергии и, при увеличении продолжительности работы более 10 минут, энергообеспечение осуществляется уже почти целиком за счет аэробных процессов.Однако, мощность аэробной системы энегообеспечения примерно в 3 раза ниже мощности фосфагенной, и в 2 раза — мощности анаэробной гликолитической системы (см. табл.)
Основные биоэнергетические характеристики метаболических процессов — источников энергии при мышечной деятельности
| Метабол. процесс |
Критерий мощности | Макс. энерго- емкость, кДж/кГ |
||
| Макс. мощность, кДж/кГ *мин |
Время достижен. макс. мощн. физ. работы, с | Время удержан. работо способ. на уровне макс. мощн., с |
||
| Алактный анаэробн. | 3770 | 2 — 3 | 6 — 8 | 630 |
| Гликолит. анаэробн. | 2500 | 15 — 20 | 90 — 250 | 1050 |
| Аэробный | 1000 | 90 — 180 | 360 — 600 | Бесконечно |
Вместе с тем, аэробный механизм ресинтеза АТФ отличается наибольшей производительностью и экономичностью. В повседневных условиях жизни на его долю приходится иногда более 90% от общего количества энергопродукции организма. В качестве субстратов окисления при этом используются все основные питательные вещества: углеводы, жиры в виде аминокислот. Вклад белков в общий объем аэробной энергопродукции очень мал. А вот углеводы и жиры используются в качестве субстратов аэробного окисления до тех пор, пока они доступны мышцам.
Аэробное расщепление углеводов до определенной стадии (до образования пировиноградной кислоты) осуществляется так же, как и при анаэробном гликолизе. Но в аэробных условиях пировиноградная кислота не превращается в молочную кислоту, а окисляется далее до углекислого газа и воды, которые легко выводятся из организма. При этом из одной глюкозной единицы гликогена в конечном итоге образуется 39 молекул АТФ. Таким образом, аэробное окисление гликогена более эффективно, чем анаэробное. Еще больше энергии выделяется при окислении жиров. В среднем 1 моль смеси различных специфических организму человека жирных кислот обеспечивает ресинтез 138 молей АТФ. При одинаковом по весу расходе гликогена и жирных кислот, последние обеспечивают почти в три раза больше энергии, чем углеводы. Жиры, таким образом, обладают наибольшей энергоемкостью из всех биоэнергетических субстратов (см. табл.)
Просмотров: 8130



